Die Retina
Die Netzhaut des Auges, die Retina, enthält als vorgeschobener Gehirnteil nicht nur Sinneszellen, sondern auch ein ausgeklügeltes neuronales Netzwerk. Wir untersuchen die Informationsverarbeitung in diesem Modellsystem des Gehirns.
Auf den ersten Blick ähnelt unser Auge einer Kamera. Optische Bausteine wie die Linse oder die Iris erzeugen ein Bild unserer Umwelt auf der Retina, die den Augenhintergrund auskleidet. Die Retina ist ein hochspezialisiertes neuronales Netzwerk, das in der Lage ist, unterschiedliche Aspekte der bildlichen Information zu extrahieren und zu verarbeiten.
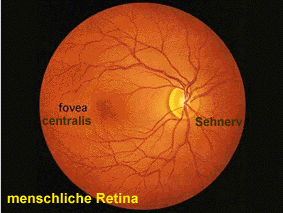
Ein Arzt, der mit einem Augenspiegel die Retina betrachtet, sieht in etwa folgendes Bild. Man kann zwei spezialisierte Regionen klar erkennen: rechts (in Richtung Nase) verlassen die Axone der retinalen Nervenzellen das Auge und bilden den optischen Nerven. An diesem „blinden Fleck“ treten auch die Blutgefäße in das Auge ein. Links (in Richtung Schläfe) erkennt man einen oval geformten, blutgefäßfreien Bereich, den „gelben Fleck“. Er enthält die Fovea, die Stelle des schärfsten Sehens.
Die Retina des Menschen ist ca. 200 – 300 µm dick. Bei allen Wirbeltieren besteht die Retina aus mehreren Schichten. Abbildung 2 zeigt einen Schnitt durch die Retina, links in Form eines Schemas, das exemplarisch einige der retinalen Zelltypen darstellt, rechts in Form eines histologischen Schnittes.

Erstaunlicherweise sind die lichtempfindlichen Photorezeptoren am weitesten vom einfallenden Licht entfernt. Das Licht muss alle retinalen Schichten durchdringen, bevor es auf die lichtempfindlichen Bereiche der Photorezeptoren trifft, die Außensegmente. Wir unterscheiden zwei Arten von Photorezeptoren. Die Stäbchen weisen eine hohe Lichtempfindlichkeit auf. Mit ihnen sehen wir bei Nacht und in der Dämmerung. Die Zapfen sind weit weniger empfindlich und erlauben uns das Sehen bei hellem Tageslicht und die Unterscheidung von Farben. Ein menschliches Auge enthält 100 – 120 Millionen Stäbchen und ca. 6 Millionen Zapfen. Die Zellkörper der Photorezeptoren bilden die äußere Körnerschicht (englisch outer nuclear layer, ONL). Die Absorption von Licht löst im Außensegment eine Kette intrazellulärer Reaktionen aus, die schließlich in einer elektrischen Antwort münden: die Membranspannung des Photorezeptors wird negativer, d.h. die Membran hyperpolarisiert und die Menge des Neurotransmitters Glutamat, der an der Synapse freigesetzt wird, nimmt ab.
In der ersten synaptischen Schicht der Retina, der äußeren plexiformen Schicht (OPL), übertragen die Photorezeptoren die Information an die Bipolarzellen und die Horizontalzellen. Bipolarzellen leiten die Information in die innere plexiforme Schicht (IPL) weiter, wo sie auf Ganglienzellen und Amakrinzellen übertragen wird. Die Ganglienzellen, von denen es ca. 1 Million pro Auge gibt, senden die Information vom Auge ins Gehirn. Der Weg vom Photorezeptor zur Ganglienzelle mutet wie ein Staffellauf an. In Wirklichkeit wird aber an jedem Kontaktpunkt das Signal neu verrechnet und gegebenenfalls verändert. So entstehen bereits in der OPL zwei ganz unterschiedliche Signale. Während Stäbchen und Zapfen bei Belichtung hyperpolarisieren, können Bipolarzellen unterschiedliche Signale erzeugen. Bei Belichtung hyperpolarisieren die „OFF-Bipolarzellen“, d.h. sie werden inhibiert. Die ON-Bipolarzellen depolarisieren, d.h. sie werden erregt. Die Unterteilung in ein ON- und ein OFF-System bleibt auch im Gehirn erhalten. Darüber hinaus wird in den synaptischen Schichten die Information, die von benachbarten Retinaorten stammt, verglichen. Ermöglicht wird das durch Zellen mit langen lateralen Fortsätzen, wie den Horizontalzellen in der OPL und den Amakrinzellen in der IPL. Das Signal, das eine Ganglienzelle ins Gehirn sendet, repräsentiert deshalb nicht einfach einen Helligkeitswert, sondern kann deutlich komplexer sein. Es wird am besten durch die Eigenschaften des rezeptiven Feldes der Zelle beschrieben. Bei vielen Ganglienzellen besteht das rezeptive Feld aus zwei konzentrisch angeordneten Regionen, die antagonistisch zueinander sind. Fällt Licht in das Zentrum einer ON-Zentrumsganglienzelle, wird sie erregt und feuert eine Salve von Aktionspotentialen. Fällt das Licht in das Umfeld, wird die Zelle gehemmt. Eine OFF-Zentrumsganglienzelle wird dagegen durch „Licht-Aus“ im Zentrum erregt, aber durch Licht-Aus im Umfeld gehemmt. Diese Ganglienzellen übermitteln Information über den Kontrast zwischen dem Zentrum und dem Umfeld, aber nicht über die wirkliche Helligkeit. Der falsche Eindruck vieler optischer Täuschungen beruht genau darauf, dass die wirkliche Helligkeit nicht an das Gehirn weitergeleitet, sondern aus der Kontrastinformation neu berechnet wird. Einige Ganglienzellen haben rezeptive Felder, die noch komplexer sind. Sie reagieren z.B. nur dann, wenn sich ein Objekt in einer bestimmten Richtung durch ihr rezeptives Feld bewegt. Wir vermuten, dass die Retina 15 verschiedene Typen retinaler Ganglienzellen enthält. Jede wirkt wie ein neuronaler Filter, der einen Aspekt visueller Information (Farbe, Bewegung, Kontrast usw.) aus dem komplexen Abbild unserer Umwelt auf unserer Retina extrahiert. Wir interessieren uns vor allem dafür, welche Rolle verschiedene Klassen von Ionenkanälen und Botenstoffen spielen, die Lichtantworten in den verschiedenen retinalen Zelltypen zu erzeugen und zu formen.

Die Retina ist ein faszinierendes zelluläres Netzwerk. Die in der Retina enthaltenen 130 – 160 Millionen Zellen können in ca. 60 verschiedene Zellklassen eingeteilt werden, Informationen werden innerhalb der Netzhaut durch Fortsätze mit einer Gesamtlänge von ca. 10 km weitergeleitet und an ca. 10 Milliarden Synapsen übertragen. Wenn Sie mehr über unsere Arbeit erfahren möchten, folgen Sie diesem Link.
Publikationen zum Thema aus unserem Institut:
Müller, F., Scholten, A., Ivanova, E., Haverkamp, S., Kremmer, E., Kaupp, U.B.
"HCN channels are differentially expressed in retinal bipolar cells and concentrated at synaptic terminals."
European Journal of Neuroscience 17 (2003) 2084-2096
Ivanova, E. and Müller, F.
"Retinal bipolar cells differ in their inventory of ion channels."
Visual Neuroscience 23 (2006) 143-154
Mataruga, A., Kremmer, E., Müller, F.
"Type 3a and 3b OFF-cone bipolar cells provide for the alternative rod pathway in the mouse retina."
Journal of Comparative Neurology 502 (2007) 1123-1137
Knop, G., Seeliger, M.W., Thiel, F., Mataruga, A., Kaupp, U.B., Friedburg, C., Tanimoto, N, Müller, F.
"Light responses in the mouse retina are prolonged upon targeted deletion of the HCN1 channel gene."
European Journal of Neuroscience 28 (2008) 2221-2230
Wässle, H., Puller, C., Müller, F., Haverkamp, S.
"Cone contacts, mosaics, and territories of bipolar cells in the mouse retina."
Journal of Neuroscience 29 (2009) 106-117