Membranen
Umschichtung von Nanopartikeln durch Lipid-Doppelschichtmembranen
Die Membranausstülpung und die Partikelaufnahme spielen eine wichtige Rolle bei der Kommunikation von Zellen mit ihrer Umgebung, beispielsweise bei der Endozytose, der Phagozytose sowie beim Eindringen von Parasiten oder Viren. Zudem erfordern verschiedene potenzielle Anwendungen von Nanopartikeln in komplexen Materialien ein besseres Verständnis ihrer zellulären Toxizität. Darüber hinaus können Nanopartikel für die gezielte Wirkstoffabgabe, zur Krebstherapie sowie als Membranbildner für biomedizinische Studien eingesetzt werden.

Wir untersuchen die passive Endozytose von Nanopartikeln mit einer Größe zwischen zehn und einigen hundert Nanometern, die von Lipid-Doppelschichtmembranen umhüllt sind. Beim Umhüllen steigt die Verformungsenergie der Membran an, während die Adhäsionsenergie aufgrund der Anziehungskräfte zwischen Partikel und Membran abnimmt. Numerische Berechnungen mit triangulierten Oberflächen ermöglichen die Ermittlung der Verformungsenergien für verschiedene Partikelformen.
Wir charakterisieren die Umhüllung von Nanopartikeln analog zu thermodynamischen Phasenübergängen. Während teilweise umhüllte Zustände bei kugelförmigen Nanopartikeln nur in Membranen mit Spannung auftreten, zeigen würfelförmige, stabförmige und ellipsoide Nanopartikel flache und tiefe teilweise umhüllte Zustände auch ohne Spannung. Hier kommt es auf die Form an! Nicht nur das Seitenverhältnis, sondern auch die lokale Krümmungsverteilung auf der Nanopartikeloberfläche steuert die Bindung und Umhüllung. Dies ist der erste entscheidende Schritt für die Wechselwirkung von Nanopartikeln mit biologischen Zellen.
Unter Druck schwankende Schalen

Eine dünne Kugelschale kann als elastische Membran betrachtet werden, die über einen Schermodul, eine Biegesteifigkeit und eine von Null verschiedene Krümmung verfügt. Im Mikrometer- und Submikrometerbereich reichen die Beispiele für solche Schalen von synthetischen hohlen Polyelektrolytkapseln mit bedeutenden technologischen Anwendungen bis hin zu biologisch relevanten Systemen wie roten Blutkörperchen und kugelförmigen Viruskapsiden. Ohne thermische Fluktuationen hängt die Stabilität elastischer Schalen gegenüber äußeren Kräften wie einem gleichmäßigen Druckfeld oder einer punktförmigen Eindrückung vom Verhältnis zwischen der Größe der Schale und der Wandstärke ab. Wie bei flachen Membranen ist zu erwarten, dass thermische Fluktuationen die mechanische Reaktion einer verformten Schale durch die Renormierung der Elastizitätskonstanten beeinflussen.
Die Schwankungen dünner Schalen unterscheiden sich jedoch qualitativ von denen flacher elastischer Membranen, was auf die Kopplung von Dehnungsmoden in der Ebene und Biegemoden durch die Krümmung zurückzuführen ist. Wir untersuchen die Verformungen dieser Schalen mithilfe von Monte-Carlo-Computersimulationen und der Störungstheorie unter Einbeziehung der Auswirkungen von Krümmung und äußerem Druck. Wir zeigen, dass thermische Schwankungen den kritischen Knickdruck verringern und das mechanische Verhalten bei punktförmigen Eindrücken abschwächen.
Durch konische Einschlüsse induzierte Knospenbildung
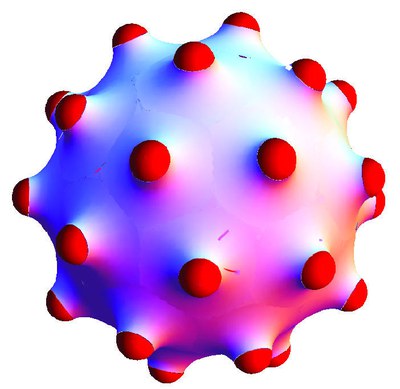
Die Knospenbildung und Vesikelbildung von Lipid-Doppelschichten ist der erste Schritt für den Stofftransport in biologischen Zellen. Die Knospenbildung kann durch gekrümmte Einschlüsse ausgelöst werden, wie beispielsweise asymmetrische Proteine oder Viren, die teilweise an der Doppelschicht haften.
Während gekrümmte Einschlüsse in einer ebenen Membran einer abstoßenden, membranvermittelten Wechselwirkung unterliegen, stellen wir eine effektive Anziehungskraft für Einschlüsse auf einer Knospenmembran fest. Diese Anziehungskraft wird durch die abgeschirmte Abstoßung der Einschlüsse auf einer gekrümmten Membran verursacht. Unter Verwendung eines Modells, das ausschließlich Biegeenergie berücksichtigt, ermitteln wir einen optimalen Knospenradius und damit eine spontane Krümmung c0 für eine gegebene Einschlussdichte, was katenoidenartigen Membranverformungsbereichen um die Einschlüsse herum entspricht. Bei der Knospenbildung aus einer Muttervesikel erzeugt die Biegeenergie allein eine Reihe entarteter Grundzustände für den Knospenradius als Funktion der Anzahl der Einschlüsse pro Knospen. Die Translationsentropie der Einschlüsse hebt diese Entartung auf, und die Minimierung der freien Energie ermöglicht dann eine eindeutige Vorhersage der Knospenradien.
Zerknittern dünner elastischer Folien

Die Verformung dünner elastischer Schichten ist ein grundlegendes Problem mit zahlreichen praktischen Anwendungen in verschiedenen physikalischen und biologischen Systemen. Zu diesen Systemen zählen makroskopische Materialien, die von dünnen Stahlplatten über dünne Gummifolien bis hin zu Papierbögen reichen, mesoskopische Materialien wie Tonplättchen, die Membran biologischer Zellen und Riesenvesikel, aber auch mikroskopische Materialien wie Viruspartikel und Kohlenstoffnanoröhren.
Mithilfe von Computersimulationen haben wir den Einfluss der Selbstvermeidung auf die Formen, die mechanischen Eigenschaften und die Faltenlängenverteilungen von zerknitterten elastischen Folien untersucht.
Verformung von Viruskapsiden

Das Genom eines Virus ist in einem schützenden Gehäuse enthalten, das als virales Kapsid bezeichnet wird. Ein virales Kapsid weist eine bemerkenswert regelmäßige Struktur auf und setzt sich aus einer festen Anzahl von Kopien eines einzigen oder weniger Arten von Kapsidproteinen zusammen. Seine Geometrie entspricht der eines Ikosaeders oder eines spiralförmigen Zylinders, es gibt jedoch auch komplexere Strukturen. Wir untersuchen die mechanischen Eigenschaften ikosaedrischer Virushüllen mittels Computersimulationen.
Wir berechnen das elastische Verhalten bei kleinen Verformungen sowie die Knickübergänge bei großen Verformungen, die sowohl stark von der Anzahl der elementaren Bausteine als auch von der Schub- und Biegeelastizität der Schale und der umgebenden Geometrie abhängen.
Mit Polymeren beschichtete Membranen
Wechselwirkungen zwischen Polymeren und Membranen spielen in vielen komplexen Systemen eine Rolle, die von den Membranen biologischer Zellen bis hin zu Mikroemulsionen reichen. So verbessert beispielsweise der Polymerzusatz in ternären Mikroemulsionen die Mischwirksamkeit des Tensids zwischen Öl und Wasser. An Membranen verankerte Polymere unterdrücken Schwankungen der Membrankonformation, d. h. der Polymerzusatz bewirkt eine effektive Versteifung der Membran.
Mithilfe von Monte-Carlo-Simulationen bestimmen wir die effektiven Krümmungs-Elastizitätskonstanten für polymerbeschichtete Membranen. Insbesondere untersuchen wir den Einfluss der Selbstvermeidung innerhalb einer einzelnen Polymerkette. Dies wurde erweitert, um auch andere Polymerarchitekturen wie Sternpolymere zu untersuchen. Mit zunehmender Armzahl der verankerten Sterne steigt die Biegesteifigkeit stark an, während der Sattelspreizmodul konstant bleibt.
Amphiphile
Amphiphile Blockcopolymere in bikontinuierlichen Mikroemulsionen

Wird einer phasengetrennten Mischung aus Öl und Wasser eine geringe Menge an Tensidmolekülen zugesetzt, bildet sich eine homogene „Mikroemulsionsphase“, die etwa gleiche Mengen an Öl und Wasser enthält. Diese Phase weist eine „bikontinuierliche“ Struktur auf, da sie aus zwei miteinander verflochtenen Netzwerken aus Öl- und Wasserkanälen besteht (siehe Abbildung).
Geringe Mengen eines amphiphilen Blockcopolymers führen zu einer dramatischen Zunahme der Öl- und Wasservolumen, die in einer bikontinuierlichen Mikroemulsion solubilisiert werden können. Der Einfluss verschiedener Blockcopolymerlängen und -konzentrationen auf die Struktur und das Phasenverhalten ternärer Mikroemulsionen wurde untersucht.
Hochpräzise Neutronenstreuungsexperimente liefern eindeutige Belege dafür, dass die Polymere gleichmäßig verteilte „pilzartige“ Konformationen auf der Tensidmembran bilden (siehe Abbildung). Auf der Grundlage dieser Beobachtungen schlagen wir einen universellen Mechanismus für das Quellverhalten vor, der auf die Veränderung der Elastizität der Membrankrümmung zurückzuführen ist.
Kubische bikontinuierliche Phasen in ternären amphiphilen Systemen

Grenzflächen in amphiphilen Systemen lassen sich oft gut durch elastische Schichten mit einer Biegesteifigkeit k, einem Sattelspreizmodul k und einer spontanen Krümmung c0 beschreiben. Die amphiphilen Monoschichten in ternären Gemischen aus Wasser und Öl können sich auf unterschiedliche Weise anordnen und so mizellare, hexagonale, lamellare sowie verschiedene dreifach periodische, bikontinuierliche kubische Phasen bilden.
Die relative Stabilität der letztgenannten Phasen lässt sich durch das Zusammenspiel ihrer universellen geometrischen Eigenschaften mit den Konzentrationsbedingungen erklären. Wir stellen fest, dass die stabilsten kubischen Phasen bei abnehmendem k<0 die einfachen und doppelten Gyroidstrukturen sind – siehe Abbildung für die minimale Gyroidfläche
Abschnüren kristalliner Bereiche in Flüssigmembranen

In einer biologischen Zelle wird die Bildung kleiner Vesikel an der Plasmamembran - ein Prozess namens Endozytose - von der Adsorption von Clathrin-Proteinen gesteuert. Diese Moleküle bilden auf der Membranoberfläche lokal ein regelmäßiges, sechseckiges Netzwerk aus. Somit entspricht die Bildung von Vesikeln aus einem solchen Bereich physikalisch dem Ausschnüren von kristallinen Bereichen, die in Flüssigmembranen eingebettet sind. Hier wird die Ausschnürung von der Linienspannung der Bereichsgrenze und von der spontanen Krümmung c0 getrieben.
Wir benutzen Monte-Carlo-Simulationen an dynamisch-triangulierten Oberflächen und Skalierungsparameter, um diesen Prozess zu untersuchen. In kristallinen Phasen entstehen bei der Bildung von Kugelschalen Gitterfehler. In einem aus Dreiecken aufgebauten hexagonalen Gitter, das in der ebenen Phase nur sechsfach koordinierte Dreieckspitzen enthält, sind dies fünf- und siebenfach koordinierte "Disklinationen" (siehe Abbildung).
Wir stellen fest, dass die Defekte an der Grenze entstehen und dann ins Innere diffundieren. Darüber hinaus ergeben unsere Simulationen: Je größer der kristalline Bereich ist, desto kleiner kann die spontane Krümmung c0 sein, damit es zu einer Ausschnürung kommt. Dieses widerspricht bisherigen Voraussagen.
Fortgeschrittene Flickerspektroskopie an flüssigen Vesikeln
Die Formen und Fluktuationen von Flüssigmembranen werden von ihrer Krümmungselastizität gesteuert, die durch zwei elastische Konstanten, die Biegesteifigkeit und die spontane Krümmung, gekennzeichnet ist. Das Spektrum der Membranfluktuationen liefert Informationen über die Werte der elastischen Konstanten.
Die Analyse von Membranfluktuationen - Flickerspektroskopie genannt - hat sich bisher auf quasikugelförmige Vesikel beschränkt. So kann zwar die Biegesteifigkeit, nicht aber die spontane Krümmung bestimmt werden. Wir haben eine neue Methode entwickelt - die fortgeschrittene Flickerspektroskopie riesiger, nicht-kugelförmiger Vesikel, die es ermöglicht, beide Parameter gleichzeitig zu messen.
Unsere Analyse basiert auf einer großen Menge Referenzdaten willkürlich triangulierter Oberflächen, die aus Monte-Carlo-Simulationen gewonnen wurden. Die Methode wurde auf die thermischen Trajektorien von Vesikelformen und auf die elastische Response von zwitterionischen Membranen auf transmembrane pH-Gradienten angewendet. Die neue Technik ermöglicht es, die Membrankrümmung als Funktion von Umgebungsbedingungen mühelos zu charakterisieren.
Wassermoleküle und Wasserstoffbrücken-Netzwerke in Bacteriorhodopsin - Molekulardynamik-Simulation vervollständigt Kristallstruktur
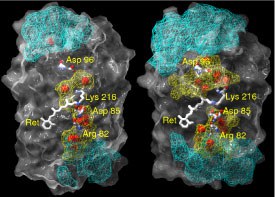
In die Struktur von Bacteriorhodpsin (bR) sind Wassermoleküle eingebaut. Die räumliche Verteilung der Wassermoleküle und das zugehörige Netzwerk aus Wasserstoffbrücken-Bindungen wurden mit Hilfe von Molekular-dynamiksimulationen in der Grundzustands-Konformation untersucht. Bei dieser Untersuchung wurde der Grundzustand gewählt, um nachzuweisen, dass sogar die Struktur mit der niedrigsten Menge an Wassermolekülen ein dichtes und stark fluktuierendes Wasserstoffbrücken-Netzwerk im fast hydrophoben Teil von Bacteriorhodopsin enthält. Das Protein wurde dazu in eine vollständig hydratisierte Lipid-Bilayer-Membran eingebettet. .
Im Vergleich zu Kristallstrukturdaten wurde in unserer Simulation eine viel höhere durchschnittliche Anzahl interner Wassermoleküle gefunden (44 statt 18). Diese Diskrepanz geht auf die hohe Beweglichkeit des H2O zwischen verschiedenen Positionen zurück. Im Durchschnitt finden wir 20 eingeschlossene und 24 diffusive Wassermoleküle im bR. Die Zeit für den Austausch aller diffusiver Wassermoleküle beträgt etwa 200 ps. Die durchschnittliche Verweilzeit eines diffusiven Wassermoleküls im Inneren von bR ist ungefähr 52 ps.
Zur Beschreibung der Wasserstoffbrücken-Netzwerke im Protein benutzen wir ein geometrisches Konstrukt, das auf dem Grotthuss-Modell beruht. So bilden sich zwischen der zytoplasmatischen Oberfläche und dem geladenen Asp96-Rest Ketten aus Wasserstoffbrücken aus. Es zeigt sich, dass die Länge einer Kette durchschnittlich aus 5 Bindungen besteht. Die durchschnittliche Lebensdauer einer ununterbrochenen Kette liegt in der Größenordnung von 0,045 ps. Die durchschnittliche Raumverteilung der Wasserstoffbrückenbindungen zwischen der zytoplasmatischen Oberfläche und Asp96 weist auf eine Art Trichter hin, der auf Asp96 gerichtet ist. Diese Untersuchung schafft eine neue Grundlage für den Protonen-Transfer über stark fluktuierende Wasserstoffbrücken-Netzwerke, um den Mechanismus des Protonenpumpens zu klären (2004).